Productivity Problems of Freshwater Warzawa Krakow 1972.
Proceedings of the IBP-UNESCO Symposim
Influence of some ecological factors on changes of the standing crop of zoobenthos of the Danube in the Bulgarian stretch
von Boris K. RUSSEV
Bulgarian Academy of Sciences, Zoological Institute,
Boul. Russki 1, Sofia, Bulgaria
(English only - most figures included)
Statistical analysis of several years of investigations on the biomass of zoobenthos in the Bulgarian stretch of the Danube shows quantitative changes of standing crop of zoobenthos and changes in the percentual presence of 16 dominating species under the influence of ecological factors. These are: consistency of the bottorn, depth, current velocity, water level, distance from shore and some other factors which change along the river or during the seasons. This proves that a general analysis of standing crop and the productivity of the Danube must take into account the above named ecological factors.
Between 1956 and 1961 and later, in 1964, 14 regular seasonal investigations on the quantitative distribution of the zoobenthos were conducted in the Bulgarian stretch of the Danube between 845 and 375 km (counting from the delta). 905 stations located in width and total length of the river sector were sampled with the help of the Petersem bottom grab (surface 0.1 m', weight 54 kg). Besides, the qualitative composition of the zoobenthos was investigated additionally in 300 stations situated a"g the Ahole Bulgarian Danube shore (Russev 1966). 240 species of benthos ha?.e been found but the frequency analysis showed that 16 species or groups am reiatively much more numerous. These are: Turbellaria ? Palaeodend~ocoehon romanodanubialis Codreanu; Nematoda, Oligochaeta ? mainly Lümw&ilus ne%wensis Michaelsen, Peloscolex velutinus (Grube), Bythonomus sp.; Gastropoda ? Theodoxus transversalis (Pfeifer), T. danubialis (Pfeifer), Lithoglyphus naticoides Pfeifer; Bivalvia ? Unio tumidus Philipsson, U. pictorum (L.), Dreissena polymorpha Pallas; Isopoda ? Jaera sarsisarsi Valk.; Gammaridae ? mainly Dikerogammarus haemobaphes fluviatilis Martinov, Chaetogammarus tenellus behningi Martinov, Pontogammarits sarsi Martinov, P. obesus Martinov and P. cravsus Martinov; Corophiic4e ? Corophium robustum Sars and C. curvispinum Sars (Amphipoda), Gomphusflavipes (Charpentier) (Odonata), Chironornidae (Diptera), l~i?drops~i?che gr. ornatula (Trichoptera).
The results of quantitative investigations. numerically and by biomass, will be treated statistically so as to present the percentual relationship of the river organisms and their standing crop. As our research vessel cannot penetrate shallows, riverain shore areas could not be treated quantitatlvely and only qualitatively?. The complete preponderance of the enumerated species or animaI groups. houever. stands. Representatives of molluses have been separated frorr thc other invertebrates serving as fish food, becuase of the weight of shells which, in lumping together, would give wrong results (S h a d i n 1948 p.438)
The substratum of the river is the most impdrtant factof deciding the distribution of benthos in the Danube. The müst favourabie habitat is mud (Corophium mud") which lies in layers over gravel. With a large development of almost all zoobenthos edible for fishes; from this substratum zoobenthes is distributed all over the river by current. Molluscs show a preference for gravel, stones, rock and slag and loam.
In comparison with this, sandy substratum offer the least favourable conditions for the zoobenthos in the Danube.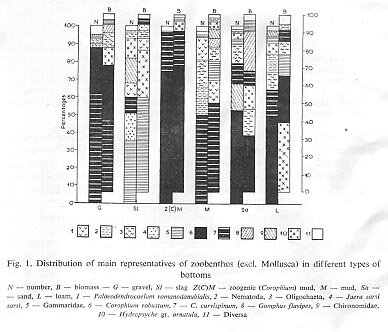
Figure 1 represents the percentual relations between the dominating species
or groups in their various kinds of substratum as to numbers and biomass, and
this was already analysed (R u s s e v 1969). Patterns of animal communities and
their distribution to ' substrata along the river's length and width have also already
been discussed (R u s s e v 1967).
Current velocity as an ecological factor should be considered in close connection with the bottom substrata as the arrangement of this is governed by the various river velocities. The pattern' of Danube sediments and substrata in relation to velocities has already been described (R u s s e v 1967, p. 33?37). It should, however, be mentioned that an increase in current velocity produces also an increased drift of organisms which can produce a sharp impoverishment of the bottom fauna. 0 1 i v a r i (1961) has contributed interesting observations on this point.
We have arranged river Velocities in three groups which relate to the distribution of dominating species: velocities up to 0.3 m,/sec., up to 0.6 m/sec. and from then upwards'. The results are given in Table 1 and Figure 2. From the data it is clear that in all three groups of river velocities, the two Corophium species are dominant, but at velocity above 0.6 m/sec. their domination is limited.
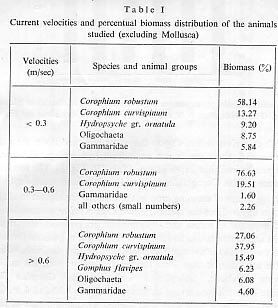
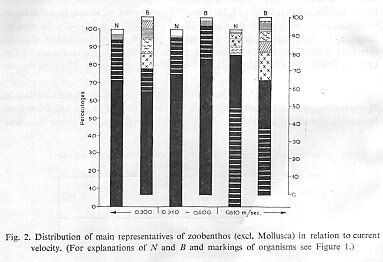
For the molluscs (Theodoxus transversalis, Lithoglyphus naticoides, Unio tumidus and others) (Tab. 11) and representatives of the Oligochaeta, lower river velocities are more favourable. A velocity of 0.3?0.6 m/sec. is characterised by the mass appearance of Palaeodendrocoelum romanodanubialis and Jaera sarsi sarsi (Isopoda) and Amphipoda. Velocities over 0.6 m/sec. show a reduction in numbers and biomass of benthos.
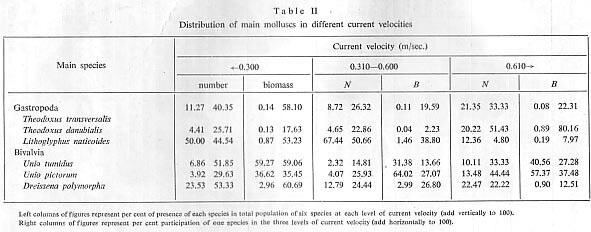
The depth of a river depends upon the water level and upon the varying relief of the bottom and, therefore, is not an independent ecological factor. We have divided our results into four depth groups (Tab. 111, Fig. 3). The results show that the dominance of the Corophiidae species in relation to river depth is evident.
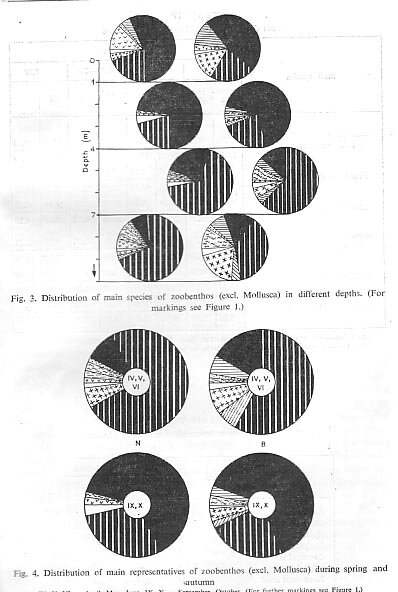
Of interest is the interdependence in the distribution of the two mass appearing species of the family Corophiidae in the Danube. Corophium robustunt dominates in number and biomass from 0 to 4 m, Corophium curvispinunt prefers depth from 4 m downwards. Such a clear distribution picture cannot be found in other dominating species.
The river level exerts a considerable influence on the quantity of zoobenthos. Therefore, investigations were carried out in times of flood, when the depth reached more than 5 m, in times of medium level between 3 and 4.6 m averagely, and for low water usually below 3 m. During flood, the general biomass of zoobenthos is only 0.755 g/m'. During niedium levels it is 1.939 and during lower levels reaches 6.351 g!m'. We believe that these differences are based on the fällowing causes:
1. During the flood the river bed is broadened and the available zoobenthos distributes itself over the whole width, causing a lowering of density per m'. During low water the reverse occurs.
2. During the flood a larger part of the bottom fauna cannot remain stationary and is carried away by the pressure of the current.
Seasonal influences are, of course, in close relationship to the water levels and the average values of standing crop in the Danube are highest in shallow waters from 0 to 1 m depth. Here the species: Palaeodendrocoelum romanodanubialis, Unio pictorum, Corophiunt robustum and Hydropsyche gr. ornatula, and representatives of Nematoda and Gammaridae find the most favourable conditions for development. With increase of depth the average values of standing crop diminish, with the lowest values in the zone deeper than 7 m. Independently, however. the mollusc species of Theodoxus danubialis, Lithoglyphus naticoides, and Goniphus flavipes and Chironomidae have a large average biomass of between 1.10 and 4 m. Oligochaeta, Dreissena and Jaera sarsi sarsi have their highest average biomass of between 4. 10 and 7 m. In depths greater than 7.10 m, Theodoxus transversalis and Unio tumidus reach their highest average biomass (Tab. IV).
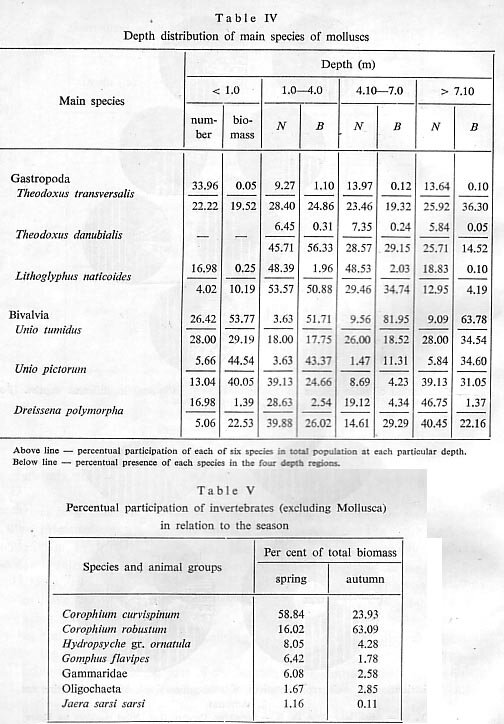
also to differences in temperature. As our investigations in the Danube were carried out mainly from April to June and in September and October, our knowledge of the relationships of dominating species or groups refers only to the spring and autumn season (Tab. V, Fig. 4).
The numbers of individuals of species of small size and weight as, e.g., Palaeodendrocoelum romanodanubialis and the two species of Corophiidae, take a larger percentage in comparison with the biomass, whereas the larger species of higher weight (Gomphus flavipes, Hydropsyche gr. ornatula and others) occupy a smaller percentage. During the autumn season the species Palaeodendrocoelum romanodanubialis, Hydropsyche gr. ornatula and exemplares from Oligochaeta, Gammaridae, Corophiidae and Chironomidae appeared in larger quantities and biomass; Nematoda, Gastropoda, Bivalvia, Jaera sarsi sarsi and Gomphus flavipes appeared somewhat in ]arge quantities during the spring. It should be mentioned that 95% of the number and biomass of Corophitim robustunz are during the autumn months, while only 55% for C. euri,ispinum.
Our results show that the average total biomass of zoobenthos in spring is 35.223 g/M2 of which molluscs are 32.485 g/M2 and the rest -2.738 g/m'. In autumn the total average biomass is 38.895 g/ml of which 34.214 g/m' are molluscs and 4.682 g/M2 the rest of the zoobenthos. The larger quantity of zoobenthos in autumn depends in our opinion upon the influence of river levels and also the development of each species during the various seasons.
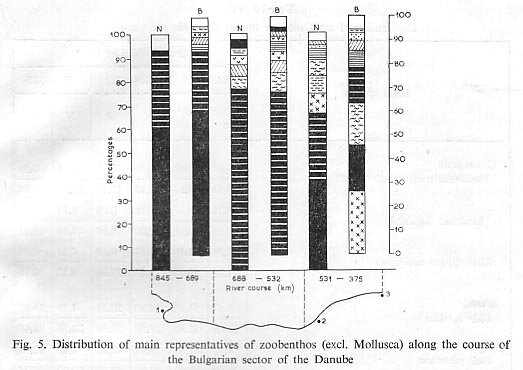
As to changes of the standing crop of zoobenthos along the Danube course, we have found that the western sector of the Bulgarian part (between 845 and 596 km of the river) is considerably richer than the eastern one (between 595 and 375 km). In the western sector we found 46.080 g/M2, in the eastern sector only 16.705 g/M2. Molluscs appeared in the western sector with 40.557 and in the eastern sector ?with 15.690 g/M2. The rest of the zoobenthos amounts to 5.523 in the western and 1.015 g/ml in the eastern part. We deduce that this is due to the lower gradient and corresponding lower current velocity, the larger percentage of gravel bottoms with lithorheophil biocenosis and the lowest participation of sandy grounds with psammorheophil biocenosis in the western sector. Also, this sector is less polluted in comparison with the eastern part where the Danube receives effluents from Bucarest and Russe. We found it expedient to group the results of percentual relationships of the dominating species or groups into three sectors of the Danube arbitrarilv chosen, but completely identical as to length. In the western sector the two Corophiidae species are H% of the biomass of invertebrate animals (without molluscs); in the middle sector their biomass is 71%, and in the eastern sector it is 35% (Tab. VI).
It seems that this more equal distribution of biomass of the dominating groups or species in the eastern sector depends upon the diminished competition of the two mass appearing species of the Corophiidae. Besides, the other ecological factors are changing, of which the main is the decrease of gravel and Corophium mud from west to east. According to our investigations (R u s s e v 1967,? p. 63) the gravel occupies in the western sector 35.16%. sand 57.42%, soft niud ? 6.04%
and loam 1.37%, whereas in the east gravel is 22.93%., sand 67.13%, soft mud 2.76% and loam 7.18%.
 The great dominance of the biomass of two Corophium species in the western sector is even more clearly visible when one considers the total arrangement of numerically dominant species in the three sectors (Fig. 5, Tab. VI).
The great dominance of the biomass of two Corophium species in the western sector is even more clearly visible when one considers the total arrangement of numerically dominant species in the three sectors (Fig. 5, Tab. VI).
Numerically prominent in the western sector are Oligochaeta, Theodoxils transversalis, Lithoglyphus naticoides, Unio tumidus and Dreissena polymorpha; in the middle sector Palaeodendrocoelut7i romanodanubiali,~" Jaera sarsi sarsi, Ganimaridae, Corophiidae and Chironomidae; in the eastern sector Theodoxus danubialis, Unio pietorum, Gomphusflai,ipes and Hydropsyche gr. ornatula (Tab. VII).
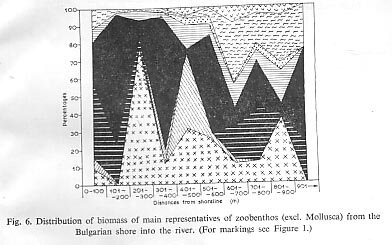
The percentual distribution of dominating species and change of the standing crop of benthos across the river has been investigated every 100 m from water edge to about 900 m distance from the Bulgarian side. The zoobenthos, excluding molluscs, is well distributed on the right shore between 0 and 200 m, where gravel occupies up to 60% of the bottom sediments and the current is at its lowest. In spite of the greater turbidity and increased current velocity, the lower temperatures and lower transparency, the region between 100 and 200 m is preferred to the more inshorezone. Changes in the % water level and the motion of the water caused by navigation makes the nearer shore zone less favourable for invertebrates.
The biomass of zoobenthos (exluding molluses) between 0 and 100 m line is
6.950 as compared with 8.814 g/ml in the zone of 100 and 200 m distance from the
shore. Further towards the river middle, the quantity of zoobenthos (excluding
molluscs) falls rapidly to 0.919 g I ~M2 between 200 and 300 m and between 300 and
500 m, first to 0.295 and then to 0.077 g~ /M2. From 500 to 900 m distance from the
shore, there is a rise again from 0.400 to 0.732 g/M2. These changing values depend
probably on the differences in current velocity and connected changes in bottom
sediments.
The average biomass of molluscs follows generally the same rules. It is highest
in the shore line between 0 and 100 and 100 and 200 m (82.697 and 40.524 g/M2);
from 200 and 300 m there is a sharp reduction to 0.779 g! IM2, frorn 300 to 400 m
the lowest value is obtained with 0.024 g/ml and from 400 to 700 m a slight. increase
is noticed and high increase between 700?800 and 800?900 m (4.293 and 4.788 g/M2).
Everywhere the zoobenthos is better developed on the right shore of the river in comparison with the left Rumanian shore. This is. in our opinion, dependent on the distribution of bottom sediments which other wise depends on the Coriolis effect, which is expressed in the physical?geographical rule of Baer?Babinet (R u s s e v 1960).
Of particular species and groups, the Nematoda, Oligochaeta, Unio tunzidus and to some degree Gomphus flavipes show a distinct presence in the shore zone between 0 and 100 m; Gammaridae, Unio pietoruni, Corophium robustum and C. curvispinum appear mostly in zone 100 to 200 m; B~vdropsyche gr. ornatula has the biggest biomass between 200 and 300 m; Theodoxus fransversalis, Dreissena polymorpha and Chironomidae between 400 and 500 m. Jaera sarsi sarsi (500?600 m), Palaeodendrocoelum romanodanubialis (600?700 m), l'heodoxus
daizubialis (700?800 m) and Lithog~vphus naticoides (800?900 m) seem to indicate
a larger biomass towards the left river shore (Tab. VIII).
Figure 6 shows the percentages of biomass occupied by the different species in the various zones.
The results obtained aid their analysis on the influences of some ecological factors upon the standing crop and changes in the percentual relationships of the dominating species show a considerable interdependence of the various factors. The optimal river conditions for zoobenthos are gravel grounds, on which a zoogenic sediment is deposed through the activity of the Corophiidae. This happens at about 100 m distance from the Bulgarian shore during relatively low current and depth of water level in a temperature of 24'C
These quantitative data can now be recalculated according to the PIB coefficient and this will be the basis for calculations of production of invertebrates in the average surface of the Danube river bed. This will allow for obtaining a general orientation in yearly average production of the zoobenthos of the Danube as depending upon the various ecological factors.
The relationships of the nutrient coefficient of the various invertebrates can be used for a possible increase of fish biomass. A similar investigation has already been made in the Danube in which the average quantitative data of zoobenthos were examined as to their possibilities to supply food to bottom living fishes (B. Russev -The zoobenthos of the Danube between 845 and 375 km as fish food. manuscript).
